Cancer rates have climbed steadily despite major advances in diagnosis, treatment, and public health initiatives. Conventional risk factors like tobacco, alcohol, and environmental exposures have been well-documented, yet a fundamental aspect of our modern environment appears to be driving this persistent increase. I believe a significant part of the answer lies in a place most cancer research has largely overlooked: the composition of your cell membranes.
I recently published a narrative review in Cureus,1 a peer-reviewed medical journal that highlights emerging perspectives in clinical science and disease prevention. In this paper, I examine how shifts in dietary fats over the last century — particularly the increase in linoleic acid (LA) intake — may have fundamentally altered the structure and function of cellular membranes in ways that promote cancer development.
This hypothesis centers on two underappreciated nutrients — pentadecanoic acid (C15:0) and choline — that have steadily declined in the modern diet. Drawing on historical patterns and biochemical evidence, I outline how restoring these foundational elements may help rebalance cellular function in ways that matter for cancer prevention, setting the stage for a different way of thinking about risk, nutrition, and long-term health.
I discuss the central findings of my paper in the sections that follow, but I encourage you to read the full version (linked below) or download the simplified version linked at the end of the article, so you can see the evidence and reasoning in full. Understanding how modern dietary patterns have altered fundamental cellular structures helps clarify why cancer risk continues to rise — and what you can do to address it at the root.

The Historical Rise of Cancer Parallels Shifts in Fat Intake
Throughout the 19th century, cancer was considered rare in clinical records and population surveys. In the United States, estimated incidence rates hovered around 60 to 80 cases per 100,000 people. These numbers remained relatively stable through the mid-1800s, partly because life expectancy was short — averaging just 35 years — and many tumors went undiagnosed due to limited tools for internal examination.2
• Advances in pathology made cancer visible and measurable — It wasn’t until the adoption of histological methods in the late 1800s that neoplastic diseases became easier to confirm and track. Neoplastic diseases refer to conditions caused by abnormal, uncontrolled cell growth that leads to tumor formation, including both benign and malignant growths.
By the turn of the 20th century, these diagnostic improvements were in place across U.S. medical schools and hospitals, setting the stage for more systematic recording of cancer incidence.
• From that point forward, cancer diagnoses began to rise steadily — By 1950, incidence had increased to about 130 to 160 per 100,000, and by 1975, it reached 400 per 100,000. This steady rise throughout the 20th century coincided with industrial expansion, increased urbanization, and a dramatic shift in the availability and composition of food.
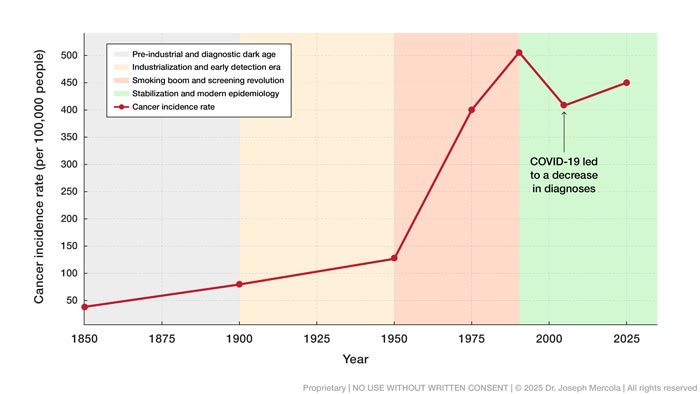
• Industrialization introduced multiple cancer-relevant exposures at once — These decades saw the mass introduction of processed foods, synthetic additives, and factory-produced seed oils into the food supply. Smoking rates also surged during this time, with about 70% of men smoking cigarettes by the early 1960s, pushing lung cancer rates upward and contributing to the overall increase in cancer burden.
Incidence peaked in 1992 at 505 per 100,000 before leveling off and slightly declining in some categories due to changes in screening practices and a reduction in tobacco use. Still, the overall trend remains markedly higher than it was in pre-industrial times.
• LA intake increased dramatically over the same timeframe — One of the most pronounced dietary shifts of the last century has been the rise in consumption of LA, an omega-6 polyunsaturated fat (PUF) that now dominates the modern fat supply. LA intake remained low through the 19th century, making up just 1% to 2% of total daily energy intake in traditional diets.
By the late 20th century, LA intake in the United States had tripled, with current estimates ranging from 7% to 10% of total energy intake. This change is reflected in human tissue data, which show that the proportion of LA stored in body fat rose from 9.1% in 1959 to 21.5% by 2008, confirming long-term biological accumulation rather than short-term dietary fluctuation.
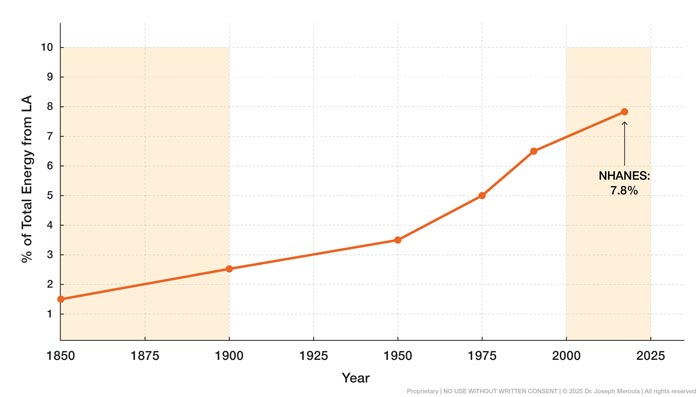
• Global comparisons show modern U.S. exposure exceeds earlier international norms — Mid-20th-century cross-population studies reported LA concentrations in blood lipids between 5.5% and 10.6% among Japanese, Nigerian, Colombian, Jamaican, and U.S. populations. Contemporary American data now exceed those levels, with current projections placing average LA exposure above 21%.
This reflects a widespread dietary shift away from animal fats toward refined vegetable oils. Oils such as safflower, sunflower, corn, and soybean now supply the majority of dietary LA, replacing traditional fats like butter, lard, and coconut oil that dominated human diets for centuries.
• Seed oils deliver vastly higher LA loads than traditional fats — Safflower oil provides more than 74 grams of LA per 100 grams of oil, sunflower oil exceeds 65 grams, and soybean oil contributes over 50 grams. By comparison, olive oil contains just under 10 grams of LA per 100 grams, while butterfat contains roughly 2.3 grams.
These figures quantify the magnitude of the dietary shift, highlighting how modern fats differ sharply from those that shaped human physiology for most of recorded history.
Changes in both the amount and type of fat consumed over the last century have reshaped the composition of human tissues, creating a sustained imbalance that warrants closer examination in relation to long-term health outcomes. But what is LA actually doing inside your cells that might explain this connection?
How LA Alters Membrane Signaling and Favors Tumor Survival
LA serves an essential biochemical role in your body, particularly in the structure and function of cellular membranes. To understand why this matters, you need to know about your mitochondria — the tiny powerhouses inside your cells that generate the energy you need to live.
These organelles depend on a specialized fat molecule called cardiolipin, found almost exclusively in the inner mitochondrial membrane. Cardiolipin acts like a scaffold that holds energy-producing protein complexes in place, and LA is the primary building block your body uses to construct it.3
• Cardiolipin integrity determines mitochondrial resilience under stress — Cardiolipin supports your mitochondria’s ability to produce energy by stabilizing protein complexes involved in oxidative phosphorylation (the process your mitochondria use to convert food into usable energy), especially during metabolic stress. The integrity of cardiolipin determines how efficiently your mitochondria operate, especially under stress.
• Excess LA is associated with disrupted inflammatory regulation and immune stress — Your immune system relies on a finely tuned inflammatory response to detect and destroy abnormal cells before they become cancerous. That process relies on brief, targeted signaling that activates immune cells, triggers programmed cell death, and then resolves.
But when LA builds up too much in your cell membranes, it changes how those signals work. Instead of turning off, the inflammation lingers. That ongoing stress makes it harder for your immune system to clear abnormal cells and creates an environment where they’re more likely to survive and grow.
• LA-rich membranes are uniquely vulnerable to oxidative damage — Because LA contains multiple double bonds, it becomes highly susceptible to oxidation. When exposed to reactive oxygen species (ROS), which are produced during normal metabolism and amplified during inflammation or toxic insult, LA becomes a target for lipid peroxidation.
Think of it like a chain reaction of rust spreading through metal. Once oxidation starts in one fatty acid, it triggers damage in neighboring molecules, releasing toxic byproducts that accumulate over time.
• This peroxidation process generates a range of reactive byproducts — This includes 4-hydroxynonenal (4-HNE), malondialdehyde (MDA), and acrolein. 4-HNE is especially harmful, as it readily forms covalent bonds with proteins through a process called adduction (permanent chemical attachment), which changes their structure and function.
These alterations interfere with mitochondrial respiration, suppress normal cell turnover, and disrupt antioxidant defenses. MDA, another product of LA oxidation, induces DNA cross-linking and mutations, while acrolein impairs protein folding and promotes chronic inflammation. Collectively, these compounds create a proinflammatory and mutagenic intracellular environment.
• LA peroxidation contributes to cancer cell survival — Aldehydes flip a master inflammatory switch called nuclear factor kappa B (NF-κB). Once activated, NF-κB tells your cells to produce inflammatory signals while simultaneously upregulating anti-apoptotic proteins such as Bcl-2 (molecules that block apoptosis or programmed cell death), making damaged cells harder to remove even when they would normally be flagged for elimination.
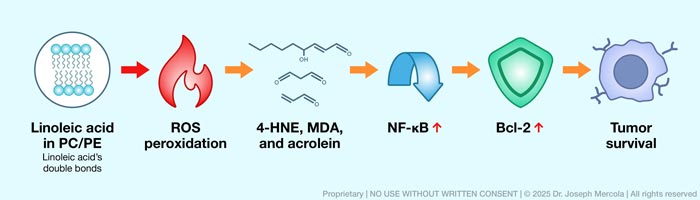
• Chronic NF-κB signaling suppresses immune effectiveness — When NF-κB stays chronically activated, inflammation persists while your immune system’s cancer-killing capacity weakens. Tumor cells exploit this state by deploying molecular “cloaking devices” — checkpoint proteins like PD-L1 that tell immune cells to stand down.
They also recruit suppressor cells that actively block immune attacks. This causes cancer cells to become invisible to the very system designed to eliminate them.
• LA may influence growth signaling in tumor cells — LA also contributes directly to cancer progression through its interaction with fatty acid–binding protein 5 (FABP5). Within tumor cells, FABP5 binds LA and enhances signaling through the mTORC1 complex, a cellular growth switch that, when chronically activated, promotes unchecked cell division.
In practical terms, this means that the LA already stored in your tissues isn’t just passive cargo — research suggests it may play a role in tumor cell growth signaling in ways that could bypass normal controls. In cancers with elevated FABP5 expression, including aggressive breast tumors, this pathway correlates with poorer outcomes, linking membrane-associated LA to sustained mitogenic drive.
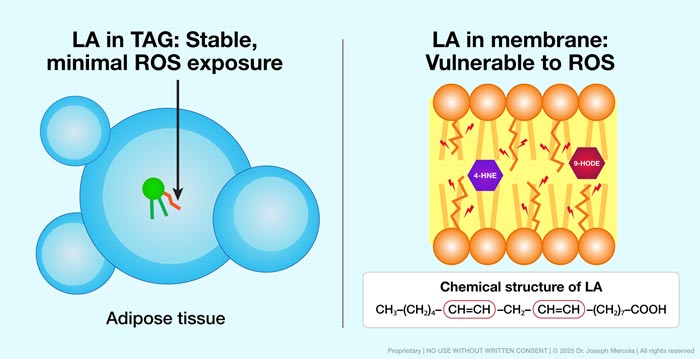
• The location of LA determines the magnitude of this risk — When stored in adipose tissue as triacylglycerol, LA remains largely inert and resistant to peroxidation. But when incorporated into phospholipid bilayers (the double-layered fat structure that forms all cell membranes), it becomes directly exposed to ROS.
In this setting, LA rapidly forms oxidized phospholipids and secondary metabolites that activate innate immune receptors and sustain inflammatory signaling. This chemistry lowers apoptotic sensitivity and reshapes immune surveillance in ways that allow tumor cells to persist.
Together, these mechanisms explain how LA supports normal cellular function under controlled conditions yet promotes immune suppression, chronic inflammation, and survival signaling when it becomes excessive. For a deeper look at how to address the chronic inflammation and metabolic disruption that shape cancer risk, read “How Metabolic Health and Inflammation Influence Cancer Risk.”
How Membrane Turnover Creates a Window of Opportunity
Once you understand how LA weakens membranes, a practical question follows naturally: How quickly can those membranes change once dietary inputs change? The answer depends on how fast your body rebuilds them, and that process moves far more quickly than most people realize.4
• Your cell membranes are not static structures — The phospholipids that make up your cellular and mitochondrial membranes are constantly broken down and rebuilt as part of normal metabolism. Studies show that about half of the phospholipids in a typical cell membrane are replaced every two to three days, with faster turnover in highly active tissues.
Liver cells renew large portions of their membranes within 24 to 48 hours, while slower-metabolizing cells such as red blood cells replace their membranes over a longer period. This ongoing renewal allows membranes to respond continuously to energy demand, oxidative stress, and nutrient availability.
• Membrane composition changes on a timescale of days, not years — Because phospholipids turn over rapidly, the fatty acids incorporated into new membranes reflect what your body has available at the time of rebuilding. Changes in dietary fat intake begin influencing membrane composition within days to weeks, rather than remaining locked in place for decades.
This helps explain why fat tissue reflects long-term dietary patterns, while membranes remain comparatively flexible and responsive. The membrane acts as the most immediate interface between what you eat and how your cells behave.
• This turnover also creates a meaningful opportunity — When you reduce dietary LA, less of it is available for membrane integration, and the signals that would otherwise emerge from its oxidation are inhibited. That structural change reduces the need to block those signals downstream.
Anti-inflammatory drugs work by interfering with signaling cascades that occur after membrane oxidation. Membrane remodeling addresses the problem earlier in the sequence — by limiting the inputs that initiate inflammatory signaling in the first place. That makes it an earlier point in the inflammatory signaling sequence.
Targeting membrane composition may offer a way to influence how your immune system perceives and responds to cellular stress, especially in tissues where LA-related oxidative stress is a concern. Among the fats that shape this response, one molecule that stands out for its stabilizing role is pentadecanoic acid (C15:0).
Pentadecanoic Acid (C15:0) — A Structurally Unique Membrane Stabilizer
Pentadecanoic acid (C15:0) is an odd-chain saturated fat that originates almost exclusively from ruminant foods, including whole-fat dairy, butter, cheese, and meat from cows, sheep, and goats. Unlike many fats that are synthesized or modified extensively within the body, C15:0 reflects direct dietary intake, which is why it has long been used as a biomarker of dairy fat consumption.5
• Modern diets have quietly depleted this essential fat — For most of human history, regular consumption of ruminant-derived foods supplied a steady background level of C15:0 that became incorporated into cellular membranes throughout the body.
As dietary patterns shifted away from full-fat dairy and pasture-raised animal foods, tissue levels of C15:0 declined alongside them. This change did not produce acute deficiency symptoms, but it altered membrane composition and their response to metabolic and inflammatory stress.
• Odd-chain structure supports membrane flexibility — What distinguishes C15:0 from more common saturated fats is its odd number of carbon atoms. Most saturated fats in modern diets have even-numbered chains that pack together in highly regular, rigid patterns within the phospholipid bilayer.
The slightly irregular structure of C15:0 disrupts this uniform packing just enough to preserve membrane flexibility while maintaining structural order. This allows membranes to bend, signal, and recover under stress without becoming disorganized or prone to damage, a balance that is especially important under metabolic or inflammatory stress.
• C15:0 resists oxidation and limits LA enrichment during membrane renewal — Because C15:0 contains no double bonds, it does not undergo lipid peroxidation. When incorporated into membrane phospholipids during normal turnover, it occupies positions that might otherwise be filled by oxidation-prone LA.
Over time, this shifts membrane composition toward greater chemical stability. Fewer vulnerable fatty acid chains embedded in membranes means fewer reactive breakdown products generated during oxidative stress, reducing pressure on nearby proteins and signaling pathways.
• C15:0 supports energy regulation through AMPK signaling — Beyond its structural role, C15:0 influences cellular metabolism by activating AMP-activated protein kinase (AMPK), a key regulator of energy balance and stress adaptation. Experimental models show increased AMPK phosphorylation (activation) following C15:0 exposure, alongside reduced mTOR signaling.
These shifts promote phospholipid remodeling patterns that favor shorter-chain and monounsaturated fatty acids, including increases in oleic acid content. The result is improved membrane flexibility and resilience under conditions such as hypoxia or nutrient stress.
• C15:0 attenuates ferroptosis risk — C15:0 has also been shown to reduce susceptibility to ferroptosis, a form of iron-dependent cell death driven by lipid peroxidation, by lowering the availability of oxidation-prone membrane lipids and stabilizing mitochondrial membranes under stress.
In the context of modern diets that overwhelm membranes with oxidation-prone fats, restoring C15:0 reintroduces a stabilizing element that human physiology evolved to rely on, particularly in tissues where membrane integrity and immune signaling play a central role in cancer risk. You can learn more about the health benefits of C15:0 in “C15:0 — Found in Dairy — May Be an Essential Fat.”
Note: Findings related to C15:0 AMPK signaling cited in this article are derived from experimental laboratory models. These findings may not directly apply to human health.
Choline’s Role in Mitochondrial and Cellular Membrane Maintenance
Alongside C15:0, another nutrient that plays an important role in membrane structure is choline. Your body uses choline to produce phosphatidylcholine (PC), one of the major phospholipids in your cell and mitochondrial membranes.
• Your cell membrane’s resilience starts with these phospholipids — PC accounts for about half of your total membrane composition, while another key membrane phospholipid, phosphatidylethanolamine (PE) adds another 20% to 30%. Both are essential for keeping membranes stable and flexible during stress, especially in tissues with high energy demands.6
• Most people don’t get enough choline from food — National survey data show that more than 90% of U.S. adults fall short of the recommended intake. Without enough choline, cells can’t produce enough PC, and the balance between PC and PE starts to shift. This weakens the structure of the membrane and reduces its ability to manage oxidative damage.
• Mitochondria rely on targeted choline transport for membrane repair — Studies have identified a specific transporter that moves choline into the inner mitochondrial membrane, where it helps build PC. When this transporter is missing or underactive, cells show increased oxidative stress, slower growth, and a shift in energy metabolism that resembles patterns seen in cancer.
• Membrane demand for choline rises under oxidative pressure — While choline is best known for its role in neurotransmitter production, most of your body’s demand for it goes toward building and repairing cell membranes. As LA accelerates membrane damage, your requirement for choline increases. Without sufficient intake, membranes remain unstable, making it easier for the effects of lipid peroxidation to spread throughout your cells.
Hence, restoring adequate choline levels may help support membrane stability and reduce the oxidative stress associated with LA-driven membrane damage, especially when combined with efforts to lower LA consumption and increase C15:0 intake.
Putting Principles Into Practice
Understanding the science only matters if it leads to action. Fortunately, you don’t need drastic measures or advanced tools. You can simply begin with the steps below, which focus on removing the main source of membrane-damaging fats and supporting a cellular environment less permissive to tumor growth:7
1. Eliminate LA sources in your diet — The largest contributor to excessive LA exposure comes from industrial seed oils, such as soybean oil, corn oil, canola oil, sunflower oil, safflower oil, and generic “vegetable oil.” These deliver LA in amounts your tissues were never designed to handle, especially when consumed daily.
Removing these oils requires more than avoiding the bottle on your counter. It also means limiting the foods made with them, including ultraprocessed foods, restaurant meals, sauces, dressings, and condiments — where these oils are often hidden.
Because of this, one of the most practical ways to reduce your exposure is to take control of the fats you use at home. Replace seed oils with options that are naturally low in LA and more stable under heat, such as grass fed butter, ghee, tallow, and coconut oil. These fats support membrane integrity rather than feeding the oxidative processes that damage it.
The goal is to reduce daily LA intake to below 5 grams, and ideally closer to 2 grams per day. Reaching that level takes consistency, especially since LA is hidden in so many everyday foods. Tracking your intake can make the process more manageable and help you stay within a range that supports healthier metabolism and immune function.
The Pax health platform includes Seed Oil Sleuth — a tool designed to estimate your LA exposure and help you monitor your intake so you can make more informed food choices over time.
2. Increase your C15:0 intake — You can raise your C15:0 levels by choosing foods naturally rich in this fat, including:
• Full-fat dairy from grass fed cows; a single cup of whole milk provides about 100 milligrams of C15:08
• Grass fed beef, which contains more C15:0 than grain-fed beef
• Lamb, especially Australian lamb
• Fish such as mullet and catfish, which have C15:0 levels comparable to milkfat
• Butter, ideally from grass fed cows
• Full-fat cheese made with animal rennet
• Organic pasture-raised eggs
• Traditional fermented dairy foods, such as kefir or cultured butter
3. Prioritize choline-rich foods — The Adequate Intake (AI) set for choline is 550 mg/day for men and 425 mg/day for women. The best food source is pastured egg yolks, but be sure to look for low-PUFA eggs.
A single egg yolk contains about 125 mg of choline, or about 23% of your daily requirement,9 making it one of the best choline sources in the American diet. Grass fed beef liver is another option that delivers a highly absorbable form of choline, with 430 mg of choline per 100-gram serving.10
4. Consider supplementation — Supplementation provides a reliable way to raise C15:0 and choline levels if your diet falls short. If you avoid animal products, getting enough choline becomes a real challenge. Plant-based sources like cruciferous vegetables offer only small amounts, and you’d need to eat unrealistic quantities to meet your needs. When diet alone isn’t enough, a supplement may be used to help fill nutritional gaps.
One underrated option is citicoline, a form of choline that’s been largely overlooked because most products use doses that are far too low to have a real effect. Research suggests effective doses may range from 500 to 2,500 milligrams (mg) per day. As for C15:0, I recommend supplementing with 250 mg of pure C15:0 daily to accelerate the restoration of healthy levels.
For additional strategies to build a cancer-resistant terrain, read “Historical Rise of Cancer and Dietary Linoleic Acid — Mechanisms and Therapeutic Strategies.” I also encourage you to download and read the simplified version of my paper to explore the evidence and mechanisms discussed here in greater detail.

Frequently Asked Questions (FAQs) About C15:0, Choline, and Cancer
Q: Why does membrane composition influence cancer risk?
A: Your cell membranes control how signals are received, how energy is produced, and how damaged cells are identified and removed. When membranes are built from fats that are chemically unstable, they become more vulnerable to oxidative damage, which disrupts these control systems. This may create an environment that favors tumor formation.
Q: How does excess LA influence cancer biology?
A: LA becomes problematic when large amounts are incorporated into cell and mitochondrial membranes. In that position, it is easily oxidized, producing reactive byproducts that disrupt signaling pathways controlling cell growth and cell death. Over time, this promotes chronic inflammation and prevents damaged cells from undergoing programmed cell death, increasing the risk that they accumulate and develop into cancer.
Q: What role does C15:0 play in stabilizing cell membranes?
A: C15:0 is a structurally stable fat that integrates into membranes without undergoing oxidation. Its odd-chain structure helps maintain membrane flexibility while preserving order, allowing membranes to function under metabolic and inflammatory stress. By occupying space that might otherwise be filled by oxidation-prone fats, C15:0 reduces the formation of damaging breakdown products and supports healthier cellular signaling.
Q: Why is choline essential for membrane repair?
A: Your body uses choline to produce the phospholipids that make up most cell and mitochondrial membranes. When choline intake is insufficient, membranes cannot be rebuilt effectively, especially after oxidative damage. This leaves cells less able to regulate signaling, manage stress, and maintain normal turnover, increasing vulnerability over time.
Q: How can I increase my C15:0 and choline intake?
A: You can help raise C15:0 levels by eating foods such as grass fed beef, whole dairy, butter, lamb, and fish like wild Alaskan salmon. Choline is found in options like pastured egg yolks and grass fed beef liver. If these foods are not regularly part of your diet, targeted supplementation may help support adequate levels and maintain membrane stability.
This article is for informational purposes only and does not constitute medical advice. Consult a qualified healthcare provider before making changes to your health regimen.
Test Your Knowledge with Today’s Quiz!
Take today’s quiz to see how much you’ve learned from yesterday’s Mercola.com article.
Which is an actual symptom of dementia?